GEN1 : DNA och Cellen Flashcards
Vad är: - cellkärnan - nukleol - mitokondreien ?
- Cellkärnan: Här lagras DNA vårt genetiska material. Består av ett yttre och ett inre membran där det yttre membranet fortsätter direkt ut i ER. Innehåller kärnporer där små molekuler kan ta sig in/ut, på detta sätt kan vissa hormoner och signalämnen komma in i kärnan och binda till receptorer i nukleoplasman och förändra gentranskription. För transport av större molekyler in hit används transportmolekyler.
- Nukleol: Är som kärnan i kärnan och har troligen en funktion vid rRNA syntes och bildningen av ribosomer • Mitokondrien är cellens energiverk, här produceras den mesta energin från näringen.
- Mitokondrien består av två biomembran, det yttre som en bubbla där det finns poriner som tillåter passage av molekyler. Det inre membranet är mycket längre och veckat för att få ut så mycket yta som möjligt, det är även mycket tätare och allt transport över det inre membranet kräver transportproteiner. Området mellan membranen kallas ofta mitokondriala matrix och här sker den oxidativa fosforyleringen där vi utvinner den mesta av kroppens energi. Mitokondrien har sitt eget DNA, RNA samt ribosomer. Dras ofta till platser i cellen där det krävs mycket energi eftersom den tillverkar det i form av ATP.
Vad är:
- Er
- Golgi
- Lysomen
- Endoplasmiskt reticulum, eller ER är den största organellen i cellen och har ett membran som sitter ihop med cellkärnan. Delar av ER har ribosomer på sig och kallas granulärt (rough) ER och vissa delar är släta (smooth) ER som bara är membran och har inga ribosomer. rER står för en stor den av cellens proteinsyntes, sER har olika funktioner i olika celler men producerar bland annat membranlipider och lagrar kalciumjoner. Det är även en viktig del av transport och sortering då proteinerna tas in i ERs lumen och packas i vesiklar som skickas vidare till Golgi. Även proteiner som tillverkas i ribosomer som ligger fritt i ER tas upp i ER genom att en signalsekvens sätts på proteinet som sedan förs till ER med hjälp av SRP (signal recognizing particle) som tas in till SRP-receptorer och för proteinet in i ERs lumen. Även olika posttranslationella modifieringar såsom glykosylering och bildandet svavelbryggor sker i ER.
- Golgiapparaten är uppdelad i cis-golgi och trans-golgi och består av en samling tillplattade membransäckar som modifierar och packar molekyler som görs i ER för att de ska kunna skickas dit de behövs. Ofta sluter golgi in det som ska skickas i vesiklar av biomembran. . Främst uppgift att sortera proteiner och skicka de till rätt plats. Märker de med olika Rab proteiner som är som adresser (RAB1). Proteiner kan även lagras i vesiklar i golgi.
- Lysosomen är en liten oregelbundet formad organell, med membran runt, som bryter ner oönskade molekyler och partiklar i cellen (främst olika cellkomponenter). Fungerar bäst vid pH runt 3.5 då dess hydrolytiska enzym (hydrolaser) har lättast att katalysera reaktioner, protonpumpar i membranet hjälper till att hålla pH lågt. Avfallsprodukterna utsöndras oftast via exocytos. Lysosomen kan släppa ut sina digestiva enzym i cellen som då genomgår autolys och äter upp sig själv.
Vad är Peroxisomen, Proteasomen och ribosomen?
- Peroxisomen är aktiv vid oxidativa reaktioner i cellen och bryter främst ner aminosyror och fettsyror. Kan bland annat vara aktiv i beta oxidationen av långa fettsyror och är även involverad i pentos-phosphate-pathway. Är en lite membranförsluten vesikel där väteperoxidas bildas av de nedbrytna fettsyrorna och omvandlar sedan det till syre och vatten med hjälp av enzymet katalas.
- Proteasomen bryter ned proteiner i cellen, kan dels vara för att de är felveckade och därmed farliga för cellen eller för att det finns en överproduktion. Den är cylinderformad med proteaserna inlåsta i mitten så att de inte skadar cellen om det inte är medvetet. Ubiqutin är ett litet protein som med hjälp av en ligas fästs på proteiner som ska brytas ner i proteosomen.
- Ribosom som är uppbyggt av två subenheter som i sin tur är uppbyggda av olika proteiner och rRNA. Det är här som proteinerna bildas utav mRNA i processen translatation. Den lilla subenheten läser av koden på mRNA och matchar ihop med dess antikodon på tRNA så att rätt aminosyra förs till polypeptiden. Den stora subenheten katalyserar peptidbindningarna mellan aminosyrorna. Den har tre stycken sites, (E, P och A).
Vad består cellmembranet av?
-
Cellmembranet är det yttre lagret som är uppbyggt främst av fosfolipider, glykolipider men har även proteiner och kolesterol i sig. Är amfifilt och skapas naturligt utav hydrofoba krafter. Reglerar vad som får komma in och ut ur cellen. Den hydrofila delen sitter alltid antingen inåt, utåt eller båda och den hydrofoba är i mitten i det ihopsittande täta membranet.
- Fosfolipider:
- Mättade eller saturerade – ger inga dubbelbindningar och blir organiserade
- Omättade eller icke saturerade – ger dubbelbindningar och blir mer rörliga
- Glykolipider:
- Fäster till en till sockergrupp
- Bara på det yttre membranet
- Viktig för transport
- Kolesterol:
- Behövs för rörelse i membranet
- Sätter sig mellan fosfolipidsvansarna/fettsyrorna
- Membranproteiner:
- Transmembranproteiner som sitter igenom membranet på båda sidor
- Perifererala proteiner som sitter på insidan och kan antingen sitta som en del i membranet eller vara bundet till en fosfolipid
- Viktiga för transport, cellsignalering, enzymer, receptorer samt fäster till cellskelettet.
- Fosfolipider:
Vad är cytoskeltte och cytopasman för något?
- Cytoskelettet består av tre sorters filament: mikrotubili (som håller allting på plats och är viktigt för transport av saker i cellen), aktinfilament (Används mycket i fagocytos och cellrörelse) och intermediär filament, som ger cellen stabilitet men inte har så mycket med rörelsen att göra. Det håller även ihop celler genom att via membranproteiner fästa vid proteinfibrer utanför cellen och på så sätt kunna binda till andra celler. Tight-junctions proteiner bildar långa kedjor runt två intilliggande plasmamembran vilket förhindrar läckage mellan cellerna. Desmosomer gör samma sak fast är mycket starkare och håller ihop med intermediär filament.
- Cytoplasman är den gelliknande substans som omger alla organeller och fyller ut cellen, som vätskan runt om allting i princip.
Vilka delar består celldelningen av?

Vad är interfasen?
- G1, S-fasen och G2.
Interfasen är viktig för cellens tillväxt under celldelningen samt DNA-replikationen, alltså att kopiera genomet. Interfasen utgör den tid det tar för cellen att duplicera dess innehåll, för att sedan kunna genom gå mitos och cytokines → ny syntetisk cell.
Vad sker i G1 och G2 fasen?
G1: Detta kallas även växt-fasen och de här cellen bygger upp med proteiner och organeller som kommer behövas under delning.HärskerocksåtransptictionavDNA è RNA. När denna ”uppbyggnad” är klar, så kommer cellen fram till en ”check Point”. Om cellen klara checken så går den vidare till fasS.OmintesåfallerdeninenG0fas,där cellen vilar.
G2: I g2 fasen kontrollerar cellen att det finns tillräckligt med proteiner och organeller för celldelning. Om inte så produceras det mer.
Vad sker i mitosen och fram till slutet på celldelningen?
- Mitos
- Profasen
- DNA kondenserar samman till kromosomer
- Det mitotiskt nätet bildas
- Centersoler polaliseras
- Kärnmebranet börjar upplöses
- Metaphase
- Kärnmembranet upplöses helt
- Delningskromosomerna helt packerade och lägger sig på ekvatorial-planet
- Mibrutibulinet i mitosiak nätet fäster vid centromerna vid delningskromosomerna
- Anaphase
- Disjunktion sker, vilket är att delningskormosomerna delas till två syster kromosomer. Sker igenom att kohesion bryts ner.
- Miktotublulinerna blev kortare, samtidigt som polerna rör sig från varandra.
- Telophase
- Kärnmembran skapas runt det nya paren kromosmer. Och Kromosomerna blir löst DNA
- Profasen
- Cytokines
- Delning av cellen
- Är contractilringar med Actin och myosinflament som klipper plasman
- G0
- Är en vilofas där
Vad bygger upp DNA?
DNA-kedjorna består av två strängar – S och ´S, med nukleotider fästa på ”ryggraden”. Nukleotiderna i genomet är cytosin (C), adenin (A), guanin (G) och thymin (T). Dessa nukleotider binder bara till varandra genom vätebindningar, med dessa nukleotidkombinationer: A = T och G ≡ C
Hur startar replikationen av DNA?
Kortet svar på hur de går till fram till vi bytter håll på allt
DNA-replikationen startar redan i G1 på flera olika ställen (replication origins) samtidigt längs DNA-kedjan, de startar på ställen där nukleotiderna ligger i en speciell ordning, oftast där det finns många A = T, (TATA-boxar) eftersom de bara har två vätebindningar emellan sig och alltså är lättare att bryta upp. Var dessa finns och där det alltså bildas origins of replications identifieras av proteinerna CDc6 och CDt1. När sedan CDt1 och CDc6 har identifierat origins of replications och fästs till DNA-kedjan så bryter proteinet helikas helix-strukturen hos DNAt. Det sätts även fast enkelsträngande proteiner SSbp (single strand binding protein) till replikationsgaffeln som då ser till att helixstrukturen hålls isär, slät och tillgänglig tills DNA-replikationen är färdig.
I origins of replications blir det sedan som en slags bubbla där DNA-kedjorna har delats av helikaset och bildar en form av replikationsgafflar. Vid replikationsgafflarna är det sedan möjligt för DNA polymeras och en RNA primer att påbörja replikationen och syntetisera en dottersträng.
Enzymet DNA polymeras kan inte sätta sig direkt på kedjan och börja syntetisera eftersom den behöver en 3’ ände för att kunna sätta på fler nukleotider. Därför finns det någonting som heter RNA primers som kan starta en helt ny polynukleotidkedja bara genom att binda ribonucleoside triphosphater (ATP, GTP, CTP) och på såt sätt starta upp replikationen. Den gör sedan en liten sträng i 5’ till 3’ riktningen och lämnar sedan 3’ sidan fri för DNA polymeraset som fästs på DNA-kedjan med hjälp av sliding clamps att komma in och fortsätta replikationen. DNA Polymeras kan bara bygga 5’ -> 3’ eftersom skulle den bygga åt andra hållet hade 5’ änden behövt varaPolymeraset använder sig nu av föräldra-strängarna (S och S´strands) som mallar för att kunna replikera en ny exakt DNA-sträng och det fungerar eftersom A = T, T = A, C = G, G =C. DNA polymeras röra sig åt ett håll på DNA-strängen (5´till 3´) eftersom DNA polymeras bara kan addera nukleotider på 3´- änden, en anledning till att den inte kan göra det åt andra hållet är eftersom det inte hade gått att proofreada så enkelt då vilket den behöver kunna göra.

Vad är lagning och leding-strand vid replikation?
Den DNA-sträng som rör sig åt ”rätt” håll kallas för leading strand, medan den som rör sig åt ”fel” håll kallas för lagging strand. Leading strand kan börja replikeras så fort ett kort RNA-kedja (primer) satts på vid replikationsgaffeln och gjort de första paren och DNA polymeras kan sedan fortsätta eftersom helikaset fortsätter att bryta upp strängarna och presentera nya nucleotider att para ihop med.
Lagging strand fungerar lite annorlunda eftersom den inte bara kan fortsätta utan behöver göra om hela processen hela tiden eftersom den hunnit ifatt svansen på leading strand/svansen på förra okazaki fragmentet. Cellen löser det genom ett enzym som kallas DNA primas som hela tiden använder ribonucleoside trifosfater för att syntetisera nya primers på ungefär 10 baser långa som ett DNA polymeras sedan kan använda för att sätta igång. Men på lagging strand funkar inte RNA som primer på samma sätt eftersom inte DNA polymeras kan ta vid på samma sätt. Sedan används ett speciellt DNA reparationssystem för att byta ut RNA primers mot DNA och sedan kommer ett DNA ligas in och binder ihop alla lagging strandsen med 3’ änden från det nya fragmentet och 5’ änden av det gamla
Efter att kedjorna har brutits av helikas och sedan tvinnats ut av SSbp (single strand binding protein) så kommer strängen ha en hög spänning, vilket gör det svårt för kedjan att återgå till helixstrukturen. Därför finns proteinet DNA topoisomeras som sänker spänningen så att kedjorna efter DNA replikationen kan återgå till helixstrukturen.

Hur beskrivs reglering av cellcyklen översiklit och vad är CDK och cykliner?
Varje fas beror på att den föregående har fungerat felfritt och avslutats korrekt. Detta kontrolleras i checkpoints. Cellcykeln har flera checkpoints varav övergången G0 è G1 och G1 è S kontrolleras extra noga
CDK och cykliner
Progression genom cellcykeln regleras mha cykliner och CDK som komplexbinder till cyklinerna. CDK-cyklin-komplexen fosforylerar målprotein, t.ex. Rb, vilket driver cellcykeln framåt. Efter ett ”pådrivningssteg” minskar cyklinnivån för den cyklinen.
Aktiviteten hos komplexen regleras av CDKIs. I cellcykeln finns övervakningsmekanismer för DNA och kromosomer som kontrollerar att inga skadade celler replikeras. Upptäcks DNA-skada kommer checkpoint-aktivering att fördröja cellcykeln och trigga reparationsmekanismer. Om skadan är för allvarlig för att reparera kommer cellen gå i apoptos/senescens.
Det finns flera familjer av CDKIs, t.ex. p21, p27, p57. Dessa är breda och det finns även en del mer selektiva.

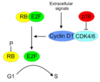
Hur reglerar RB cellcykeln?
- När cellen tar emot signaler utifrån att den ska dela så ökar cyklin D i koncentration
- När cyklin D ökar i koncentration så associerar den med ett annat protein: cyklinberoende kinas (Cdk)
- Detta Cdk-komplex fosforylerar Rb-proteinet
- När Rb-proteiner fosforyleras så släpper den transkriptionsfaktorn E2F
- När E2F blir fri stimulerar den transkription av gener/ proteiner som är viktiga att driva cellcykeln. Exempelvis:
a. Cyklin A och B: båda dessa cykliner som driver cellen genom cellcykeln
b. E2F stimulerar även cyklin E, som med ett annat cyklin-beroende-kinas (Cdk) hjälper till att fosforylera Rb - Transkriptionsfaktorer för enzymer till DNA-syntesen
- Man får en väldigt snabb och effektiv övergång från G1 till S-fasen
- S-fas → proliferation
Hur reglerar P53 cellcykeln?
- ATM aktiveras (sensorprotein som känner av hur cellen mår
- ATM fosforylerar p53 så att de släpper ifrån MDM2 (vilket i normala fall bidrar till att ubiqenin kan binda in. P53 gör även mdm2 som håller den kontrollerad)
- P53 med fosfat kan binda till DNA och verka som transkriptionsfaktor för:
- P21
- DNA-reparation
- Apoptos via BAX
- Cell-vila
- p53 stödjer cellcykelstopp (arrest) för att möjliggöra DNA-reparation eller hantera cellstressen/ hypoxi
- Om cellen löser sina problem: då släpper fosforyleringen av ATR så att p53 inte fosforyleras och därav stabiliseras
- P21 ger CDK inhibering genom att hämma Cyklin D och CDK 4/6
- CDK inhibering gör så RB inte fosforyleras
- Vi får igen E2F som verkar som TF è
- Vi kommer inte gå över i S-fasen
Vilka tre nivåer av DNA uppbyggnad/ihoprullning finns?
Man brukar säga att det finns tre sycken nivåer av chromatin strukturer
- DNA viras runt histon proteinerna och bildar såkallade nucleosomer (beads on a string). Detta kallas också euchromatin och är det DNA som kan läsas av och är i sin okompakta form)
- Massor av histoner viras ihop till ett 30 nanometer tjock fiber som består av nucleosomer och ligger väldigt tätt och detta DNA kan i denna form inte läsas av i gentranskriptionen (heterchromatin).
- En ännu högre nivå av DNA packetering av den 30 nm tjocka fibertråden som sker när kromosomerna går in a metafasen under mitosen
Vad är telomeras och telmomerer?
Telomerer: Längst ut på DNAsekvenserna så sitter det telomerer som fungerar som en buffertzon vid replikationen. Eftersom lagging strand aldrig kommer kunna kopiera hela genomet finns det alltid en bit som blir över och med repeterade celldelningar kommer denna buffertzon sakta men säkert att försvinna vilket man tror är en stor anledning till att cellen då slutar att dela sig. Dessa nukleotidsekvenser är icke kodande för proteiner och repetitiva i sekvensen GGGTTA om och om igen (ungefär 1000 gånger i varje telomer). Genomet kallas alla generna som finns i kromosomerna tillsammans, medan fenotypen är alla de gener som faktiskt uttrycks (telomererna är alltså med i genomet men inte i fenotypen).
Det finns ett enzym som heter telomeras som bygger på den här telomererna efter varje celldelning. Den kommer dock oftast att bygga på lite för lite vilket gör att för varje celldelning så minskar telomererna ändå. När de till slut når hayflickgränsen efter ca 50-60 delningar (beror mycket på celltyp) så kommer cellen känna av att den har för korta telomerer och därför kommer att behöva klippa av gener i fenotypen vilket den inte vill.
Cancerceller har oftast ett överaktivt telomeras som klarar av att bygga på tillräckligt med nukleotidsekvenser på genomet att det aldrig når hayflickgränsen och kan då i princip dela sig föralltid och kan på ett sätt ses som odödliga celler.

vad är RNA?
vilka tre sorter finns?
RNA är enkelsträngat och linjärt och uppbyggt utav ribos, en fosfatgrupp och en kvävebas. Kvävebaserna kan vara adenin, guanin, cytosin eller uracil. Byggs ihop av en ryggrad av fosfodiester bindningar men är alltså inte en dubbelhelix så finns inte så mycket vätebindningar mellan kvävebaserna som det gör i DNA.
Man brukar säga att det det finns tre stora sorter av RNA.
- mRNA (messenger) görs vid transkription, och är mallen som skickas till ribosomen för att bygga proteinet. Har ett antikodon från DNA som sedan används i translationen i ribosomerna.
- rRNA (ribosomalt) fungerar som ett enzym i ribosomen och är en av byggstenarna i ribosomen.
- tRNA (transfer) är format ungefär som en treklöver där ”botten” har tre baser utåt som i translationen matchar sitt kodon till mRNAs kodon (vilket är DNA:ts antikodon). Den har då en aminosyra bundet till sitt huvud och vilken aminosyra beror på vilket kodon den har i botten och bygger då på polypeptidkedjan och vi skapar våra proteiner.
hur blir ett DNA till pre-mRNA
Från DNA till pre-mRNA – Transkriptionen
Första steget för att skapa ett protein är transkriptionen. Den går ut på att kopiera DNA till mRNA. Detta börjar alltid vid en sk promotor eller TATA-box (med TATAAAA i följd). RNA-polymeras II öppnar DNA-kedjan och syntetiserar en komplementär sträng mRNA, eftersom det alltid byggs 5’-3’ läser alltså RNA-polymeras av strängen som är 3’-5’. Den sträng som inte läses av blir alltså coding strand för den har samma kvävebassekvens som mRNAt. RNA-polymeras II fortsätter transkribera och bygga mRNA tills den stöter på ett terminus, en specifik sekvens med kvävebaser som det känner igen och släpper sedan. DNA sluter sig precis bakom RNA-pol hela tiden. mRNAt får även en poly-A-svans (massa adenin) som skyddar den från nedbrytning i 3’-änden och en cap i 5’änden som hjälper mRNat att ta sig ur nukleus. mRNAt som bildas vid transkriptionen är en direkt kopia av DNAt och är inte riktigt färdigt för att translateras till aminosyror ännu.
Hur går vi från pre-mrna till mRNA?
Från pre-mRNA till mRNA – splicing
Pre-mRNAt som är en direkt kopia av DNA består av både introner och exoner, där introner inte kodar för proteinet (men har andra funktioner som är lite oklara) och exoner är det man faktiskt är ute efter. Intronerna måste alltså klippas bort innan mRNAt kan transkriberas. Detta görs i nukleus av spliceosomen (enzymkomplex med flera subenheter bestående av snRNP, small nuclear ribonucleicproteins) som hittar GU i pre-mRNAt och binder in. Mha ATP kan spliceosomen ”vika över” mRNAt så att GU klipps av och binder till ett A som alltid sitter en bit längre ner på intronen. Ännu lite längre fram klipps pre-mRNAt igen vid AG och de två exon-ändarna sätts ihop, intronen viker upp sig igen och bryts snabbt ner. Kvar finns bara exonerna vårt pre-mRNA har blivit färdigt mRNA som kan transporteras ut i cytosolen och translateras.
Alternativ splicing ger större variation bland våra proteiner, alltså att spliceosomen klipper bort även en exon.
Hur blir mRNA till protein?
Translation sker i 3 delar:
- Initation
- Elongation
- Termination
Initation
- Iniations factor (små protein) hjälper mRNA att hitta ”small subunit”. Small binder in till 5’ på mRNA
- Small rör sig längs med (emot 3’) tills den hittar ”start codon” vilket är sekvensen 5’-AUG-3’.
- Start codon signalerar till tRNA att hämta aminosyran metioni (met) vilken är komplement till AUG.
- Met fäster vid AUG (vilket är P-site)
- è Large Subunit fäster nu in vid Met.
Elongation:
- För att ribosomen ska röra på sig så måste ett nytt t-RNA binda in vid A-site.
- När P och A-site är fyllda gör enzymet ”peptidy transferase att peptid bidningar mellan de två amiosyrorna bildas
- Amiosyran på P-site släppet sedan è bildar en kedja ifrån A-site
- Translocation
- Ribsomen flyttas 3 kvävbaser i 5’è3’ riktning. Detta gör att t-RNA:t som innan satt på P hamnar på E-site. Vid E-site så tas de bort av ribosomen.
- Detta sker om och om och bildar en kedja av aminosyror.
Termination:
- Det finns en stop-kod som signalera slut
- Sker när UAA, UGA eler UAG läses av.
- Ett protein (release faktor protein) binder till A-site
- Detta gör att kedjan brytes av och ribosom-komplexet släppet ifrån mRNA.
Kvar står vi med en amiosyrkedja è PROTEIN
Vad är metylering av DNA? även acetylering.
Metylering av DNA görs av metyltransferaser och innebär att en metylgrupp (Ch3) adderas på 5-kolet i cytosin eller 6-kolet i adenin. Samtidigt sker en deacetylering som gör att histonerna packas mer kompakt och tillsammans gör detta att genen blir svårare att läsa av. Hela kromosomer kan också packas ihop väldigt tätt så att generna i den inte kan uttryckas. Detta sker t ex med den andra x-kromosomen hos kvinnor som då blir en barr body (slumpmässigt vilken som packas ihop i olika celler i kroppen, det blir som en mosaik). Det bildar då heterochromatin som inte är läsbart istället för euchromatin.
DNA kan även acetyleras vilket gör det mer läsbart eftersom histonerna tappar sin positiva laddning, vilket minskar interaktionerna den har med de negativt laddade phosphatgrupperna på DNAt.
Vad definera en stamcell och vilka olika finsn?
Två egenskaper definierar stamcellen:
- Oändlig potential till celldelningar och samtidigt behålla dottercellerna odifferentierade
- Kapacitet att mogna ut till en eller flera olika celltyper
Potensgraden anger till vilken grad stamcellen kan differentiera.
Totipotenta stamceller är de allra tidigaste cellerna i embryot, vilka har förmågan att bilda vilken celltyp som helst i organismen inklusive celler som bildar moderkakan (placenta).
Pluripotenta stamceller som kan ge upphov till alla celler i en vuxen individ. Kan ej ge upphov till placenta.
Multipotenta stamceller ger enbart upphov till en närbesläktad familj av celler, t.ex. hematopoetiska celler blir blodceller.
Unipotenta stamceller kan endast ge upphov till en enda differentierad celltyp.
