Lecture Notes/Slides for Final Flashcards
II. The Visual System: The retina
A. The retina consists of several thin layers of cells distributed across the inside of the eye.
B. The fovea is the portion of the retina where light falls from an object that you are looking directly at. It is the portion of the retina with the highest acuity, the ability to resolve fine detail and patterns of light. Note: Acuity and sensitivity are different.
C. The optic disk is the retinal location where axons from a type of retinal cell collect and exit the eye and form the optic nerve. This is the blind spot because there are no photoreceptors in the optic disk.
D. Below is a diagram depicting the eye and how a visual image is “mapped” onto the retina:
E. A neuron’s receptive field is the location in the environment (or the surface of the body) from which an appropriate stimulus will change that cell’s activity. For example light at ‘A’ (the tip of the flame in the diagram above) will affect the activity of retinal cell in location ‘a’ in the retina. Cells in different locations in the retina have receptive fields in different locations in the visual field.
F. There are five cell types in the retina.
- Photoreceptors – The first stage in the visual system.
a. Photoreceptors - the only cell type in the visual system that is directly sensitive to light.
b. There are two types of photoreceptors:
1) Rods
a) There are about 120 million rods in the human retina.
b) Rods are highly sensitive to light and are responsible for vision in very dim
light.
c) Rods are bleached in bright light and thus unresponsive in bright light.
d) Rods are not responsible for high acuity vision (not good for fine detail). e) Rods are achromatic (insensitive to colors).
f) Rods only exist outside of the fovea.
2) Cones
a) There are about 6 million cones in the human retina.
b) Cones are less sensitive to light intensity and are inoperative in dim light.
c) Cones are sensitive to color. There are three subtypes, selectively sensitive to
red, blue, and green wavelengths of light. d) Cones are most concentrated in the fovea.
c. Photoreceptors project to the bipolar cells. 2. Bipolar cells (BPs) - Retinal ganglion cells (RGCs)
a. RGCs are the only output cell type in the retina.
b. RGCs are the only means by which information from the eye gets to the rest of the
visual system and their axons form the optic tract. 4. Horizontal cells (HCs) - Amacrine cells (ACs)
G. Why is it that only the RGCs have axons?
1. Axons are needed for long-distance transfer of information. In the retina, the cells are very close together and so don’t need action potentials or axons. Also, communication by PSPs may be able to convey information that is more subtle than can be conveyed by the AP frequency code (recall that PSPs are graded).
2. Only the RGCs and ACs generate action potentials. The rest of the cell types use graded depolarization to release neurotransmitter to the next cell. A depolarization
increases neurotransmitter release. Small depolarizations cause small release of neurotransmitter; large depolarizations cause large release of neurotransmitter.
H. The relationship between different cell types in the retina.
- The retina is “inside-out” with the photoreceptors furthest away from the light (at the very back of the eye) and the RGCs the closest to the light. Thus light must pass through the other cell types to reach the photoreceptors.
- This works because all the cells in the eye, except the photoreceptors, are translucent.
- Also, at the foveal pit all cell types, except the photoreceptors, are pushed out of the way (see 9.15).

The retina consists of:
several thin layers of cells distributed across the inside of the eye.
What is the fovea?
The fovea is the portion of the retina where light falls from an object that you are looking directly at. It is the portion of the retina with the highest acuity, the ability to resolve fine detail and patterns of light. Note: Acuity and sensitivity are different.
What is the optic disc?
The optic disk is the retinal location where axons from a type of retinal cell collect and exit the eye and form the optic nerve. This is the blind spot because there are no photoreceptors in the optic disk.
diagram depicting the eye and how a visual image is “mapped” onto the retina:

What is a neuron’s receptive field?
A neuron’s receptive field is the location in the environment (or the surface of the body) from which an appropriate stimulus will change that cell’s activity. For example light at ‘A’ (the tip of the flame in the diagram above) will affect the activity of retinal cell in location ‘a’ in the retina. Cells in different locations in the retina have receptive fields in different locations in the visual field.
What are the five cell types in the retina?
Photoreceptors, bipolar cells, retinal ganglion cells, horizontal cells, and amacrine cells

Photoreceptors – The first stage in the visual system.
a. Photoreceptors - the only cell type in the visual system that is directly sensitive to light.
b. There are two types of photoreceptors:
1) Rods
a) There are about 120 million rods in the human retina.
b) Rods are highly sensitive to light and are responsible for vision in very dim
light.
c) Rods are bleached in bright light and thus unresponsive in bright light.
d) Rods are not responsible for high acuity vision (not good for fine detail). e) Rods are achromatic (insensitive to colors).
f) Rods only exist outside of the fovea.
2) Cones
a) There are about 6 million cones in the human retina.
b) Cones are less sensitive to light intensity and are inoperative in dim light.
c) Cones are sensitive to color. There are three subtypes, selectively sensitive to
red, blue, and green wavelengths of light. d) Cones are most concentrated in the fovea.
c. Photoreceptors project to the bipolar cells.

Retinal ganglion cells (RGCs)
a. RGCs are the only output cell type in the retina.
b. RGCs are the only means by which information from the eye gets to the rest of the
visual system and their axons form the optic tract.

Why is it that only the RGCs have axons?
- Axons are needed for long-distance transfer of information. In the retina, the cells are very close together and so don’t need action potentials or axons. Also, communication by PSPs may be able to convey information that is more subtle than can be conveyed by the AP frequency code (recall that PSPs are graded).
- Only the RGCs and ACs generate action potentials. The rest of the cell types use graded depolarization to release neurotransmitter to the next cell. A depolarization
increases neurotransmitter release. Small depolarizations cause small release of neurotransmitter; large depolarizations cause large release of neurotransmitter.
The relationship between different cell types in the retina.
- The retina is “inside-out” with the photoreceptors furthest away from the light (at the very back of the eye) and the RGCs the closest to the light. Thus light must pass through the other cell types to reach the photoreceptors.
- This works because all the cells in the eye, except the photoreceptors, are translucent.
- Also, at the foveal pit all cell types, except the photoreceptors, are pushed out of the way (see 9.15).
What is phototransduction?
Phototransduction is how light energy leads to a change in membrane potential.
Below is a diagram depicting how light changes photoreceptor membrane potential.
- The resting membrane potential of photoreceptors is -30 mV and this is in the dark.
- The maximum hyperpolarization is down to -65 mV and this is produced by bright light.
- Glutamate is the neurotransmitter used by photoreceptors. The greater the intensity of light, the less neurotransmitter released.

How does light produce the graded hyperpolarization?
A. The ligand-gated Na+ channels in the outer segment membrane are open in the dark, causing depolarization (to the “resting membrane potential” of -30 mV).
B. These ligand-gated channels are like receptors, but they are “inside-out”, meaning that they bind their ligand cGMP to a binding site on the intracellular face of the Na+ channel and this opens the channel.
How does light decrease the concentration of cGMP?
- The photopigment, which is highly concentrated in the membrane of the disks in the outer segment of the photoreceptors, is purple in the dark. When it absorbs light the photopigment is bleached to a pale yellow.
- The photopigment is called rhodopsin, and it consists of two parts…
a. Opsin is a protein.
b. Retinal, which is the only light sensitive molecule anywhere in the visual system.
The precursor of retinal is vitamin A. Retinal exists in 2 conformations:
1) In the dark it is 11-cis-retinal.
2) A photon of light will switch it to the all trans-retinal conformation.
- The steps in cGMP activation are as follows:
a. Opsin passes through the membrane seven times (i.e. it is a metabotropic or G- protein-coupled receptor).
b. The release of retinal from opsin allows opsin to change shape and this activates a G-protein (transducin).
c. The G-protein (G) dissociates and travels along the membrane and activates an enzyme (cGMP phosphodiesterase).
d. cGMP phosphodiesterase converts cGMP to GMP, and thus lowers the concentration of cGMP.
e. In the dark, cGMP is bound to the Na+ channel. Light decreases the concentration of cGMP, causing cGMP to disassociate from the channel. Consequently, the channels close and the photoreceptor cells hyperpolarize.

Why do we have this type of system (what is the advantage)?
a. The increased surface area and increased photopigment produced by having the photopigment molecules on the stacked disks, instead of on the Na+ channels, increases the chance of the light being detected by a rod. This system is so sensitive that a single photon can produce a detectable change in membrane potential of a rod type photoreceptor.
b. The use of G-proteins allows for amplification; each molecule of opsin can activate many G-proteins, each of which, in turn, can activate many enzymes of cGMP phosphodiesterase, each of which can, in turn, convert many molecules of cGMP into GMP.

A comparison of events triggered by G-protein-neurotransmitter receptor and photopigment

Rods versus Cones
A. Rods contain rhodopsin and are more light sensitive than cones (a single photon of light
may be detectable by rods but not by cones).
1. One reason for this is that the process of signal amplification is greater in rods.
2. Cones have coneopsin instead of rhodopsin.
a. There are three different types of coneopsin: red, green, and blue. Meaning that
they are most sensitive to light in the red, or green or blue wavelengths.
b. The difference between the three forms of coneopsin is small. The difference is a small change in the amino acid sequence allowing for maximal sensitivity to different wavelengths of light. As an interesting tidbit, there are 2 versions of the red coneopsin gene in men and these produce red opsins that vary by a single amino acid. This difference results in the two red coneopsins being maximally sensitive to slightly different wavelengths of red light. Thus, men with different versions of this opsin do not perceive the same thing when they see a red object. This answers the philosophical question about whether different people see objects, colors etc. in the same way. They don’t.
3. Another reason that rods are more sensitive to light is that the outer segment of rods is larger; therefore they have a larger surface area to absorb light.
4. Rods also have more photopigment densely packed into the membrane of the optic disks, so they absorb more light.
B. In bright light the photopigment in rods (but not cones) is saturated (bleached). Therefore rods aren’t functional in bright light, while cones are.

1. This means that we have two parallel visual systems, one for bright light and one for very dim light.
2. At night colors appear to be muted. But the same spectral frequencies exist in bright and dim light. The reason we don’t perceive colors in dim light is that cones don’t work in dim light, so we don’t perceive color because we’re using a part of the visual system that is color blind.
Below is a diagram exemplifying receptive fields of RGCs:

What is a receptive field?
A neuron’s receptive field is the location in the environment (or the surface of the body) from which an appropriate stimulus will change that cell’s activity. The term Receptive Field applies to cells in the visual system other that RGCs and also applies to neurons in other sensory modalities (e.g., touch, auditory).
Some features of receptive fields of RGCs:
- The receptive fields of RGCs are circular.
- The receptive fields of RGCs vary in size.
- Receptive fields of adjoining RGCs may overlap.
What are the mechanisms that account for the differences in size of receptive fields?
- RGCs on the edges of the retina (so their receptive fields are in the periphery of the visual field) collect information from a greater number of photoreceptors than do RGCs closer to the fovea. Therefore, their receptive fields are larger.
- Convergence is a situation where many neurons converge onto a few neurons. (There are about 120-130 million photoreceptors and only 1 million RGCs; therefore there is a lot of convergence of photoreceptors onto RGCs).
This is shown in the diagram below:

Convergence of synaptic input to RGC = large receptive field. Mostly in periphery of retina.
No convergence = small receptive field Mostly in fovea. - Divergence is few (or one unit) projecting onto many units. Divergence: One or a few units projecting to many units.
4. In the fovea there is less convergence of photoreceptors onto RGC (via bipolar cells) than in the periphery of the retina and this is a mechanism that explains the differences in size of the receptive fields of RGCs.

Divergence (image)

Light Sensitivity and Convergence
Convergence also helps to explain light sensitivity. There is less convergence in the cone system, therefore RGCs receiving input from cones are not as sensitive to light.
Receptive fields overlap and the consequence of this is that light from one point in the visual field will affect many different RGCs. This is depicted in the diagram below:

Receptive fields of retinal cells are modeled as concentric circles.
- There are two types of concentric fields, on center and off center (for both BPs and RGCs).
- These two RGCs are defined by their response to light in the center of their receptive fields. On center RGCs are turned on (generate AP) by light and Off Center RGCs are turned off by light in there receptive field centers.
- Off center RGCs generate more action potential generation in the dark than do on center RGCs.
- These concentric fields have antagonistic centers and surrounds because light falling in the center of their receptive fields has the opposite effect of light in their receptive field surrounds.
- Below is a diagram showing an on and off center RGC:

Center/surround properties.
- Antagonistic… effect of light on the center is opposite to the effect of light on the surround, and if light covers both the center and the surround, the effects cancel out.
- That our visual system is set up with centers and surrounds that are antagonistic to each other suggests that the visual system is more sensitive to contrasts in the intensity of illumination than to total brightness.
- See the PowerPoint figure showing the responses of RGCs with different receptive fields just outside versus just inside the border of the light reflected off of a sheet of white paper on a black background..
a. For an On Center RGC, when the light completely fills the center and only part of the surround, the net effect is an increase in action potentials.
b. At a corner the response is greater because the center is still completely filled and less of the surround is filled.
c. For an On Center RGC, light falling only on the receptive field surround actually suppresses the ongoing rate of AP production.
d. When light either fills both the center and the surround or it is dark there is little to no net effect on action potential production. The action potential rate is nearly the same as the baseline condition. - On center and off center RGCs overlap completely in the retina and their response to light is opposite to each other.
Why is the visual system organized to detect contrasts?
- The logical answer is that contrasts in light intensity are more informative than the overall illumination. Where are the contrasts in the figure below?
- Another way to say this is that the amount of light reflected by an object can vary dramatically depending upon ambient illumination, but we do not want our perceptions to vary dramatically in different lighting conditions.
Mechanisms for on center versus off center receptive field properties.
3: Glutamate in inhibitory here because a decrease in neurotransmitter release results in a depolarization.
- It is because of a difference in receptor type expressed by BP cells. The receptor expressed by the On Center BP cell produces an IPSP in response to glutamate and the Off Center BP cell produces an EPSP in response to glutamate.
- When is a neurotransmitter excitatory? Answer: When its presence produces depolarization.
Refer to numbers in diagram above:
#1: Glutamate, which is a transmitter released by the bipolar cells, is excitatory here because an increase in neurotransmitter release results in a depolarization.
#2: Glutamate is excitatory here because a decrease in neurotransmitter release results in a hyperpolarization.
(A useful rule of thumb: If the direction of polarization (“synaptic sign”) is the same in two neurons where one is being driven by the other, then the neurotransmitter between them is excitatory. In contrast, if the direction of polarization is opposite, the neurotransmitter is inhibitory. Also, remember the general rule that neurotransmitter release by a presynaptic terminal is increased by depolarization and decreased by hyperpolarization).
- So, the same neurotransmitter (glutamate) is released onto both ON and OFF center BP cells but there are different receptors for that neurotransmitter the BP cells (glutamate inhibits the receptor expressed by the ON center BP cell and glutamate excites the receptor on the OFF center BP cell) and this accounts for the difference between on and off center receptive fields.
- Why do we have both ON and OFF center systems? The on center system is most sensitive to increases in illumination and off center system is most sensitive to decreases in illumination.


Mechanisms for the formation of an antagonistic surround
A. The key mechanism in the formation of the antagonistic surround is the horizontal cell.
B. The diagram below shows the role of a horizontal cell (HC) when light falls on a photoreceptor (PR) in a bipolar cell’s (BP) and RGC’s surround:
C. How does light falling on the center and surround of a BC or RGC receptive field produce antagonistic effects? Light in the surround of a bipolar cell’s receptive fieldproduces the opposite effect on the membrane potential of a photoreceptor than light falling in the center. This is because of the horizontal cell’s synaptic connection between photoreceptors in the surround and photoreceptors in the center of a bipolar cell’s and retinal ganglion cell’s receptive field.
D. Figure 9.22 in Bear et al. is an excellent depiction of a single bipolar cell, with synaptic inputs from photoreceptors from its center, surround, and outside of its receptive field.

What happens to the output from the retina? (Where do axons of retinal ganglion cells (RGCs) go?)

overview conception of the organization of the visual systems

Different portions of the visual system (or visual systems) have different functions. For example:
- The superior colliculus is a tracking system for orienting the eyes towards visual (and also auditory and somatosensory) stimuli in the environment.
- The suprachiasmatic nucleus of the hypothalamus (SCn) is like the
brain’s clock. Neurons in the SCn have intrinsic firing frequencies of about 24 hours. Things such as sleep/wake cycles are controlled by the SCn. Light input to the SCn from the retina resets the SCn clock to the rising and the setting of the sun.
What is the superior colliculus?
- The superior colliculus is a tracking system for orienting the eyes towards visual (and also auditory and somatosensory) stimuli in the environment.
What is the suprachiasmatic nucleus?
The suprachiasmatic nucleus of the hypothalamus (SCn) is like the
brain’s clock. Neurons in the SCn have intrinsic firing frequencies of about 24 hours. Things such as sleep/wake cycles are controlled by the SCn. Light input to the SCn from the retina resets the SCn clock to the rising and the setting of the sun.
What is a Visuotopic map?
Visuotopic map: every point in the visual world is mapped in a point to point fashion onto various brain areas in the visual system (including the retina).
What is a Retinotopic map?
every point in the retina in mapped in a point to point fashion onto various brain areas in the visual system.
Visuotopic and retinotopic maps are essentially the same. What is different is whether the visual field or the retina is used as the initial point of reference. The visual world is repeatedly represented by visuotopic maps in different brain regions (to date there are 25-30 known visuotopic maps in the brain!!).
The diagram below shows how the retinas are divided into halves at the fovea and how the RGCs from the temporal hemiretinas project to the ipsilateral (same side) of the brain while the nasal hemiretinas project to the contralateral (opposite) side of the brain:
- The retinas are vertically divided in half through the fovea.
- The two halves of the retina have different patterns of projections:
a. Nasal hemiretinas project to the opposite side of the brain (contralateral projections).
b. Temporal hemiretinas project to the same side of the brain (ipsilateral projections).

Below is a diagram depicting how a visuotopic map is produced. However, the general answer is that the orderly representations of the visual field in the brain are produced by orderly patterns of anatomical connections.
- This diagram represents a bird’s eye view.
- Light from objects in the extreme periphery of the visual field (A and F)
to the contralateral eyes is blocked by the nose, so they are detected only in the ipsilateral eye. - The superior colliculus on each side of the brain contains a complete visual representation of the contralateral visual field.
- So, the mechanism for the formation of the visuotopic map is that the
nasal hemiretina’s RGCs project in an orderly manner to the contralateral side of the brain and the temporal hemiretina’s RGCs project in an orderly manner to the ipsilateral side of the brain. Also, the axons from RGCs in either eye that are activated by light from the same object converge on the same neurons in the superior colliculus. - Therefore, if you were to record from neurons at point B in the superior colliculus, the cells there could be driven by inputs from the right eye, the left eye, or both eyes.
- At F or A locations in the superior colliculus the cells respond to light input only from the contralateral hemiretina.

Below is a diagram depicting how information from different sensory modalities is mapped in the superior colliculus:
- There are several functionally distinct layers in the superior colliculus, and cells in each layer respond to different sensory modalities.
a. Cells in the visual layer of the superior colliculus respond to visual information from a single location. Neurons the auditory layer of the superior colliculus respond to auditory from a single location, and cells in the somatosensory layer of the superior colliculus respond to touch from a single location. These modality specific maps are aligned with each other. For example, a visual stimulus at location B (in the visual field) will cause cells in location B in the superior colliculus to fire. If that visual stimulus makes a noise (like a train whistle) then that will drive neurons at location B (in the auditory field).
b. Stimulating neurons in location B of the motor layer of the superior colliculus will cause the eyes and head to move to look towards location B in the visual field. - So one way we integrate information from different modalities is that we bring the information together in the brain.

There are two types of receptive fields - On-center and Off-center, because there are two types of RGCs: On center and off center

On and Off-center receptive fields have antagonistic center-surround properties (image)
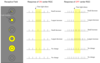
How Contrasts in Light Intensity affect RGC With Center-Surround Receptive Fields

Mechanism of On- vs. Off-center RGC receptive fields (image)

Mechanism of antagonistic center-surround property of RGC receptive fields (image)

Important functional consequences of RGC projections
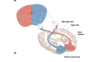
Visual field diagram

The role of the superior colliculus in the visual system
A. The superior colliculus acts as a tracking mechanism; it points the eyes and head toward a visual object.
B. Below is a diagram depicting the superior colliculus and its inputs from the retina:
- Neurons in the anterior portion of the superior colliculus respond to a visual image on the center of gaze from both eyes (but only from the contralateral visual field).
- Neurons in the posterior portion of the superior colliculus respond to a visual image in the periphery of the visual field on the contralateral side.
- Neurons in the center portion (at B) in the superior colliculus receive synaptic input from retinal ganglion cells (RGCs) from both eyes that respond to light at position B in the contralateral visual field.
4. This anatomical pattern of connections is the mechanism for generation of retinotopic maps in the superior colliculus. (See Figure 10.7).

Below is a cross section diagram of the superior colliculus showing how visual, auditory, somatosensory and motor maps are aligned with each other:
- Neurons in different layers in the superior colliculus respond to input from different sensory modalities: visual, auditory, and somatosensory.
- Each layer contains a map of a particular sensory modality, and these maps are all aligned (or in register) with each other.
- The motor layer contains motor neurons, not sensory neurons. Stimulating neurons at position B in the motor layer of the superior colliculus causes the head and eyes to move to look towards location B in the visual field.
a. Axons from the motor layer of the superior colliculus terminate in the oculomotor nucleus and neurons from the oculomotor nucleus project to muscles that move the eye.
b. Axons from the motor layer of the superior colliculus also terminate onto motor neurons which project to muscles of the neck.

Blindsight
- Blindsight is “sight” that remains when a person is cortically blind. (The eyes and optic nerve work properly, but the visual cortex is damaged so that the person cannot perceive visual images).
- A person that is cortically blind cannot “see” or identify visual objects, but can localize and track objects in visual space and they do this without awareness.
- The superior colliculus is homologous to the optic tectum, which is the main visual brain area in non-mammalian vertebrates. Apparently, our superior colliculus became specialized for tracking and lost the ability to contribute to object identification, as that function expanded in visual cortex.
Below is a diagram of a side view of a human brain depicting some of the major pathways in the cortical subsystem of the visual system in a simplified form:

Neurons in different regions of the visual cortex (V) have different functions.
a. Neurons in V5 are highly sensitive to movement but not color of objects in the visual field.
b. Neurons in V4 are highly sensitive to color but not movement of objects in the visual field.
The diagram below shows the pattern of nasal and temporal hemiretina projections to the 6 layers of the lateral geniculate nucleus of the thalamus (LGN):
- The basic form of projections from the retinal ganglion cells to the LGN is similar to that of the retinal ganglion cells to the superior colliculus, so that neurons in the LGN respond only to visual images in the contralateral visual field.
 - The LGN has six layers and each layer is retinotopically mapped, with neurons in each layer receiving input from the RGCs from only one hemiretina. Consequently, the neurons in the LGN are monocularly (respond to input from one eye) driven. Each of these maps are aligned with each other.
- Why this repetition of visuotopic maps in the LGN? The cells in the different layers are processing different aspects of the visual image.
- There are physical differences in the cells of the different layers:
a. The cells in layers 1 and 2 are larger and are called magnocellular. Cells here respond to movement and are achromatic (do not respond to color).
b. The cells in layers 3 through 6 are smaller and are called parvocellular. Cells here are color-sensitive and are more sensitive to detail.
This is an example of parallel processing. The idea is that different aspects of a visual object (aspects like color, movement, shape) are coded by different parallel pathways. - Projections to the primary visual cortex (V1) from the LGN are solely ipsilateral. Therefore, neurons in V1 respond exclusively to visual input from the contralateral visual field. This means that if the right V1 is destroyed, you would be blind to input in the left visual field (but still have “blindsight” due to an intact superior colliculus).


The diagram below illustrates the visuotopic/retinotopic map in primary visual cortex:
- Although there is a one-to-one relationship between locations in the visual field and locations in the cortical areas that encode them, almost 1/2 of the neurons in V1 code for the fovea region even though this is only a very tiny portion of the visual field. Apparently, this is because it takes a lot of neural machinery to decode the complex and detailed information that we can perceive from this area of highest visual acuity.

The visual field divided into quadrants

How do you recognize something as a face?
- Visual processing occurs in stages.
- Retina processing followed by V1 processing followed by V2, V3 processing….
In V1 (primary visual cortex) how is information analyzed?
- The visual field is broken up into approximately 1000 parts and information from each
location is analyzed by neurons in ~ 1000 different cortical modules. The diagram
below shows both the visuotopic/retinotopic map in V1 and how it is parceled into cortical modules:

Cortical modules
A. Cortical modules are hypothetical constructs but they have a physiological basis. Conceptually, each cortical module in primary visual cortex can be thought of as a computer containing all of the neural machinery needed for the initial analysis of the visual information in a single location in the visual field. Different neurons in each cortical module analyze different aspects of a visual stimulus. Some (e.g., in “blobs”) analyze the color of an object, while other neurons (e.g., those in “Orientation Columns”) analyze the shape or form of an object.
B. Some features of cortical modules:
- There are about 1000 of them in primary visual cortex with adjacent cortical modules analyzing inputs from adjacent locations in the visual field (forming a visuotopic “map” of the world onto primary visual cortex).
- They are cubes (2 mm x 2 mm x 2 mm).
- There are about 300,000 neurons in each cortical module.
- A cortical module contains 4 ocular dominance columns (actually shaped like slabs).
- A cortical module has 36 orientation columns (also shaped like slabs).
- A cortical module has 16 blobs (ironically, these are shaped like columns).
C. Figure 10.26 shows a diagrammed example of a cortical module. Neurons in a single cortical module all respond to visual input from the same particular location in the visual field. That is, their receptive fields all have similar locations. Understand Figure 10.26 1. What defines an ocular dominance column is that all the neurons in it respond primarily and preferentially to input from the same eye. Neurons in layer 4C respond to input from only one eye (one hemiretina), while the neurons in the layers above and below are driven by input from both eyes (i.e., the ipsilateral temporal hemiretina and contralateral nasal hemiretina), with the input from one hemiretina having a dominant influence. 2. Cells in blobs are color sensitive but insensitive to shape. There are 16 blobs in each cortical module, 4 per ocular dominance column. Neurons in Blobs project mainly to V4, which is specialized for color perception. 3. Information about the form of an object arises mostly from cells in the orientation column, and information about the color of an object arises mostly from cell in the blobs. Somehow this is integrated.
Below is a diagram depicting a cortical module with neurons and the stimuli that drives them:

What drives the activity of neurons in an orientation column (i.e. what is their adequate stimulus?)
- A spot of light or a spot of darkness will only weakly drive these neurons.
- An elongated stimulus is what drives these neurons (a bar of light or dark of a particular orientation).
- All the neurons in a particular orientation column respond to a line of the same particular orientation.
- Neurons in adjacent orientation columns respond to lines with a 10  difference in rotation. Thus there is a progressive rotation of line orientation that drives these neurons as you move across the columns. If you record activity across all 36 orientation columns there will be a complete 360   rotation of the orientation of the “best stimulus.”
- Your perception on a visual stimulus is thought to depend upon which subsets of cells within the relevant cortical modules are active.
What are the mechanisms generating orientation columns?
- Neurons in orientation columns have antagonistic center surround receptive fields, but they are not circular, rather they are elongated.
- Below is a diagram of two so called “simple cell” receptive fields from two neurons in different orientation columns in V1 (they can be either on- or off-center).
- These neurons are most sensitive to bars that fill their center without impinging on the surround. They aren’t sensitive to spots of light because spots don’t fill much of the long center.
- Why are these neurons sensitive to orientation of a bar of light? They are sensitive to orientation because if the light stimulus isn’t aligned with the receptive field center, then the light stimulus will fall on parts of both the center and the antagonistic surround.
- This specific type of receptive field is called a “simple cell” receptive field.
- For comparison, there are also cells that have complex receptive fields, in which the stimulus can be anywhere within the receptive field, but it still must have a particular orientation. (Cells with simple receptive field response properties are most concentrated close to layer four in V1. Cells with “complex” receptive fields are most concentrated further above and below layer four and cells with even more complex receptive fields are concentrated even further above and below layer four).
- How do these receptive fields arise? They are believed to arise because these neurons receive highly organized patterns of input from neurons from the previous stage of processing. For example, a “simple cell” that specifically responds to a vertical bar of light is thought to receive inputs from a set of neurons each of which has a circular center-surround organization with the centers of the input neurons’ receptive fields being aligned vertically with each other.

The inferotemporal cortex (IT)
A. The IT cortex has subsets of neurons that respond selectively to highly complex visual stimuli (hands, faces, food, etc.)
B. Three questions to keep in mind:
1. How do our perceptions of stimuli in different categories (hands vs. faces vs. chairs) arise?
2. How do we do this despite the fact that the image of the same object can be very different (open vs. closed book or frontal vs. profile view of a face)?
3. How do you recognize different individuals within a category (one book vs. another and one face vs. another)?
Cytoarchitecture and projections to and from striate cortex

The on “complex cell” receptive field

3 parallel pathways of visual information

How do we recognize objects as belonging to a particular category?
Answer: There are specific neurons (object recognition neurons) in the IT that respond only to specific objects within a particular category (i.e. hand, faces, chairs). Researchers have recorded from neurons in IT cortex of a monkey while projecting images on a screen that the subject can see.
- It was found that some IT neurons responded only to a particular class of images.
 - They found some neurons that responded only to faces. These neurons increased their rate of action potential production (increased their firing rate) whenever the image of a face was on the screen but not when a hand or a book or any other object was shown on the screen.
- The possibility was tested that these neurons might actually be responding to particular portion of the face and not to the face as a whole. When these researches blanked out particular portions of the face (for example the eyes or mouth) the neurons still responded to that image with increases in firing rate. However, when the face was scrambled, the neurons no longer responded.
- Both of these findings suggest that these are face recognition neurons, not neurons simply responding to the eye or mouth or some other subpart of the face. The above evidence also suggests that it is the spatial relationship among the subfeatures of the face that is important in inducing the neuron to respond.
How do we continue to recognize objects as the same even if the image of the object changes (i.e. different orientation of a face or a book)?
- As shown in the figure below, a face was shown that was tilted to different degrees (e.g., 0 vs. 45 vs. 90 degrees). It was found that the face recognition neurons still responded to all of the different tilts with about the same rate of action potential production. Note that changing the tilt of the object doesn’t change the spatial relationship among the features that make up the face.
These neurons also respond with similar numbers of action potentials to large and small faces. Note that the size of the face also doesn’t affect the spatial relationship among its features.
- But what about views that dramatically change the type of image that an object casts onto the retina (for example, frontal versus profile)? Results show that a neuron that responds to a frontal view of a face responds less and less as the face is rotated to a profile view, an orientation to which it does not respond. Other neurons in IT respond to a profile view of a face but not to the frontal view.
So, it is thought that our perception of object consistency is produced by the integration of information from neurons that respond to different views of the same object.

How do we recognize particular individuals within a category?
- There is a cross-neuron pattern of responses which is believed to be the equivalent of the “face-signature” of a particular individual. As exemplified in the figure below, one particular face produces a particular cross-neuron pattern of activity and a different face produces a different across-neuron pattern of activity in the subset of face responsive neurons in IT.

The above evidence is all correlative and “correlation doesn’t prove causation” so other evidence is needed. The correlative evidence argues that neurons in IT cortex produce the perception of a face, but it doesn’t demonstrate that. There is also some lesion data that supports the hypothesis.
A. Widespread bilateral damage of IT cortex produces a profound deficit in the ability to recognize and categorize objects, across categories. This supports the idea that IT cortex is involved in object recognition.
B. Prosopagnosia (literally means face [prosop], failure to know [agnosia]), is a syndrome in humans.
- They suffer a profound disability in recognizing faces (even pictures of themselves, spouse, their children).
- It is caused by small bilateral lesions in a particular portion of IT, or lesions of neural inputs to IT cortex.
 - The patient may have no other visual deficits (can read, recognize chairs, etc) and can also recognize people by their voices.
C. This lesion data is consistent with the electrophysiological evidence. So, there is converging evidence for IT neurons being object recognition neurons.
Face cells firing frequency changes with repeated exposures - learning new faces

Below is a side view of a cat brain and spinal cord (with the legs also).
- Walking, trotting and galloping are all different patterns of locomotion, produced by different contraction patterns of muscles. Where are the mechanisms responsible for producing locomotion? If you were to cut across the anterior portion of the midbrain (mesencephalic cats)…
a. The animals are paralyzed and can’t consciously move their lower body.
b. The cat’s body is supported in a sling and its legs are placed on a treadmill.
c. If the treadmill is turned on at a slow speed the “paralyzed” animal slowly walks with nearly perfect limb coordination and foot placement.
d. If the treadmill is turned up to a higher speed the cat trots faster with the proper coordination.
e. If the treadmill is turned up to an even higher speed the cat gallops with the proper coordination.
f. So, even though the brain is dissociated from the spinal cord, the animal still produces several forms of well coordinated locomotion.
The conclusion is that the neural mechanisms necessary to produce several forms of locomotion are in the brainstem and spinal cord and that input from the forebrain is not necessary.

Central Pattern Generator
- The symbol for a CPG is … This represents 3 neurons (A, B, C) firing in a cycling pattern.
- A CPG is a network of neurons that produces rhythmic output by its anatomical and synaptic interactions added to the intrinsic properties of its individual neurons. (spontaneously active neuron(s)). A CPG can generate a complex patterned output even if the CPG is isolated from any patterned input.

Below is a diagrammatic example of the firing pattern produced by a CPG:

Command Neuron
- A command neuron is a single neuron that generates a complex behavior by its
synaptic effects on the neurons to which it projects. - Below is an example of a command neuron:
- The different synaptic effects produced by the command neuron can generate different patterns of activity in the -motor neurons ( -MN), thus generating a patterned behavior. Note that even though this command neuron secretes one NT, different effects are produced in the recipient cells due to different receptors for the NT.

Command Group
A command group is a set of neurons with similar properties that together generate a complex behavior. That is, a command group does that same thing as a command neuron but consists of multiple neurons.
What evidence suggests that mechanisms for locomotion reside in the brainstem and spinal cord and not in the forebrain?
The figure below diagrams the main components of the neurocircuitry generating locomotion:
- Evidence from “mesencephalic” animals suggests that mechanisms for locomotion reside in the brainstem and spinal cord. A mesencephalic animal is an animal with a cut at the anterior portion of the midbrain, which will separate the forebrain from the brainstem and spinal cord. These animals are paralyzed and can’t consciously control their body.
- The mesencephalic animal is put in a sling to support its body weight and its legs are placed on a treadmill. When the treadmill is turned on…
a. at low treadmill speeds, the animal will walk with the appropriate coordination. b. at a higher speed, the animal will trot with the appropriate coordination.
c. at an even higher speed, the animal will gallop with the appropriate coordination. - Next the mesencephalic animal is put in a sling to support its body weight with its legs hanging in the air. If you electrically stimulate in an area in the midbrain called the mesencephalic locomotor nucleus (MLn)…
a. With low intensity stimulation the mesencephalic animal’s legs will move in a walking pattern.
b. With medium intensity stimulation the animal’s legs will move in a trotting pattern.
c. With high intensity stimulation the animal’s legs will move in a galloping pattern.
- All of this evidence with mesencephalic animals suggests that the neural mechanisms sufficient to produce patterned locomotion reside in the brainstem and spinal cord and not in the forebrain.

What are the mechanisms generating locomotion?
- There are multiple CPGs in the spinal cord and the neurons of the CPGs project to (alpha)-MNs.
- The fundamental rhythm of locomotion is produced by these spinal CPGs.
What turns the CPGs on in mesencephalic animals?
- When the treadmill is turned on sensory feedback from the moving legs turns on the CPGs.
a. Proprioception is sensory information that provides information about the location of parts of your body. One of the several types of sensory receptors that provide different types of proprioceptive information is the “muscle spindles.” These are embedded in muscles and provide information about muscle length.
b. There are also different sensory organs embedded in ligaments that tell about the angle of rotation of a joint and other sensory organs embedded in tendons that provided information about muscle tension. All are proprioceptors and their combined information is integrated, largely without awareness, to provide information about the location of our body parts.
c. So, when the treadmill is turned on, the legs are dragged backwards which changes the sensory feedback information from the muscles spindles and other proprioceptors and this information is sent to the spinal cord and this switches on the CPGs, and patterned movement is generated. Proprioceptive information can also regulate the rate of CPG activity. - There is a command neuron (command group) projection from the mesencephalic locomotor nucleus that synapses onto CPGs in the spinal cord.
a. You can remove sensory feedback information to the spinal cord and stimulate in the mesencephalic locomotor nucleus and you can still get coordinated movement (although not as well coordinated as when there is sensory feedback information involved).
b. In the “normal” animal there are neurons in the forebrain that project to command neurons in the mesencephalic locomotor nucleus and this is the presumed mechanism for the voluntary initiation of locomotion.
What evidence suggests that there are multiple CPGs?
- If you take a mesencephalic animal and transect (cut) the spinal cord (between the lumbar and cervical enlargements) as shown in the diagram on page 3 and then put the animal on the treadmill the animal will still generate a locomotion pattern.
- The front legs will move in alternation with each other and the back legs will move in alternation with each other, but forelimbs and hindlimbs no longer move in correct coordination with each other. This suggests that there is at least one CPG that generates the rhythm produced by the front legs and another CPG that generates the movement produced by the back legs.
How do you get coordinated activity between CPGs?
There are “coordinating fibers” that connect the CPGs together.
Below is a diagram of a cross section through the spinal cord:
- There is an anatomical separation of sensory input (which enters the dorsal horn) and motor output (which exits the ventral horn).
- There are many sensory organs in skin, muscle, tendons, ligaments etc. providing information about: heat, cold, light touch, pressure, pain, muscle length, muscle tension, joint angle, etc. (the somatosensory system). The cell bodies of the neurons carrying this sensory information from the sensory receptors are all in the dorsal root ganglion. (A ganglion is a collection of neuron cell bodies in the PNS. The same structure in the CNS is called a nucleus).
- Some of this sensory input projects to spinal CPGs (and to various other areas in the brainstem) from where it is relayed from the thalamus to sensory cortex.

How many CPGs are there? Are there two CPGs or many CPGs?
- There are many CPGs and they are distributed throughout the spinal cord, particularly in the cervical and lumbar enlargements.
- The evidence for this is if you take a thin section from anywhere in the spinal cord and put it in a dish, patterned output from the alpha motor neurons can be generated.
- Below is a diagram depicting a thin slice of spinal cord in a dish. We are recording from the left and right ventral roots and stimulating one of the dorsal roots (or adding glutamate to the bath).
- So, you can get patterned motor output which suggests that the mechanisms for producing patterned output exist within even a thin slice of spinal cord. This can be done with sections throughout the spinal cord. Therefore, there are multiple CPGs in the spinal cord and their activity can be initiated by sensory stimulation (dorsal root stimulation).
- The CPGs can also be turned on by adding glutamate into the bath. That is, glutamate will generate rhythmic ventral root activity.
- Now bisect the thin section of spinal cord. Afterwards, you can still get rhythmic motor output from the left and the right sides but now the output from the two sides is no longer coordinated with each other.
a. Therefore, there are CPGs in both the left and right sides of the spinal cord.
b. Why was the activity from the left and right sides coordinated before the bisection? Answer: because of “coordinating fibers” connecting the CPGs together.

Below is a diagram depicting the spinal cord and the inputs onto the spinal cord.

Inputs to CPGs that can trigger (turn on) CPGs and can modulate their rates.
- Coordinating fibers
- Sensory input
- Command input (from the mesencephalic locomotor nucleus).
The mechanisms of locomotion are somatotopically organized.
- There is an anterior - posterior somatotopic organization: CPGs towards the front of the spinal cord control the front legs and sensory information from the front legs synapse onto these CPGs. Conversely, CPGs towards the posterior/caudal end of the spinal cord control the back legs and they receive sensory information from the back legs.
- There is a left/right somatotopic organization: CPGs on the left half of the spinal cord synapse onto -motor neurons ( -mn) in the left half of the spinal cord and these synapse onto muscles in the left half of the body (there are both longitudinal and transverse coordinating fibers).
What kind of organization do locomotion mechanisms have?
Locomotion mechanisms have a modular organization. There are separate modules (e.g., each CPG with its inputs and outputs could be considered a module) that can operate independently but only works correctly when all the modules are connected together. Also, the system can work to some extent without sensory feedback, or without command input, or with the coordinating fibers cut.
What are the benefits of modular organization?
Why does this type of organization (modular organization) exist? This type of organization allows for a lot of flexibility and adaptability. If one component is taken out, the rest of the system will still work. Also, the sensory feedback to CPGs can allow the CPGs to adapt to different conditions (e.g., walking uphill or on ice) without higher levels needing to work out every detailed aspect of the movements required under the existing circumstances.
Are these mechanisms (modular organization etc.) just used for producing locomotion?
The answer is probably no.
- These mechanisms produce more than one type of rhythm: walking, trotting, galloping. Locomotion is a very complex process that has lots of different patterns of activity.
- Interconnecting fibers allow for the flexibility of sequences of the patterns one can get.
- Swimming, flying, walking with two or four legs, all use the same mechanisms.
- It is possible that these same mechanisms are used for any type of rhythmic output of the legs, including learned behaviors like riding a bicycle. The question of general importance in neuroscience is: Are neural circuits dedicated to one behavior or can they be used for different behaviors?
a. There is evidence that the neurocircuits that are used in locomotion are also used for scratching (evidence from Scott Currie’s lab at UCR).
b. So, maybe some neural circuits are not just dedicated to the production of any single behavior but rather are involved in controlling multiple behaviors.
Central Pattern Generators (CPGs)…a specific example.
A. Below is a concrete example of a CPG. This is taken from the Syed et al. article in the Reader.
B. The question asked in this article is: What is the nature of a CPG? How do neurons interact to produce a rhythmic output?
C. What they did to answer this question was to isolate the neurons thought to be involved in breathing in the pond snail and let them regrow in a Petri dish.
D. What does this CPG do in the pond snail?
1. The pond snail lives in water, but breaths air.
2. The pond snail comes to the surface and does a few cycles of inhalation/exhalation
and this is via a CPG (There are three neurons in this CPG).
E. Figure #1 of the paper shows the nervous system of the pond snail. It has no brain, just distributed ganglia. The three neurons of the CPG are in three different ganglia.
F. Syed et al. removed these three neurons out of the pond snail’s nervous system and kept them alive in a Petri dish. The neurons reconnected within 24 hours.
G. Below is a diagram depicting the three neurons of the pond snail respiratory CPG and their synaptic connections.
- All of the synaptic connections between these three neurons are inhibitory except for two, one of which is excitatory and the other is mixed, producing first inhibition followed by excitation.
- You wouldn’t want to breathe in and out at the same time. So IP3 and VD4, which produce exhalation and inhalation, respectively, have reciprocal inhibition and thus cannot be active at the same time.
- DA1 releases dopamine and is spontaneously active.
- IP3 and VD4 are not spontaneously active (and we don’t know what neurotransmitter they release).
H. Evidence for this wiring diagram.
- How do you know that these three neurons, by themselves, comprise the CPG? Are these three neurons sufficient to generate the patterned output? (Figure #3 of the Syed et al article).
a. They extracted the three neurons from the nervous system of the pond snail and put them in a Petri dish. They recorded simultaneously from all three of the neurons and compared this with recordings of the same neurons (from a different pond snail) in the intact snail.
b. Syed et al. found that the neurons could generate the respiratory rhythm in culture. Thus, these three neurons are sufficient to produce the rhythm. - Does it take all three neurons to generate the respiratory rhythm or only one or two?
a. They cultured each neuron in isolation and recorded from them.
b. They did not get the respiratory rhythm, suggesting that one neuron is not sufficient to generate the respiratory rhythm.
c. IP3 and VD4 were silent.
d. DA1 was spontaneously active, but it did not fire action potentials rhythmically. 3. Does it take all three neurons to generate the respiratory rhythm or only two?
a. They cultured every possible pair of the neurons.
b. They did not get the respiratory rhythm with any pair of neurons, suggesting that two neurons are not sufficient to generate the respiratory rhythm and that all three are needed. - What are the synaptic connections of these three neurons?
a. Again, they cultured every possible pair of neurons.
b. IP3 and VD4…(figure #2C of Syed et al article). When they stimulated IP3 and recorded VD4 they got an IPSP in VD4. When they stimulated VD4 andrecorded IP3 they got and IPSP in IP3. So these neurons have reciprocal inhibitory connections.
c. DA1 and VD4…(figure #2A of Syed et al article). When they stimulated DA1 and recorded in VD4 they got an IPSP in VD4, so DA1 has an Inhibitory synapse onto VD4.
d. DA1 and IP3… (Fig. 2B of the Syed article). When they stimulated DA1 and recorded from IP3 they got a hyperpolarization followed by action potentials, so DA1 has a mixed synapse onto IP3 and this produces a complex PSP, consisting of a fast IPSP followed by a slow EPSP.
I. What are the synaptic interactions that produce the pattern?
- The pond snail lives under the water but breaths air. The respiratory circuit is somehow inhibited when the pond snail is under the water.
- The pond snail swims up to the surface and when the “mouth” breaks the surface of the water, the pond snail starts to breathe. When the “mouth” breaks the surface there is a release of the inhibition on the respiratory circuit (unknown mechanism) but likely by a command neuron of some kind.
- Because of the release of inhibition, DA1 is able to begin firing spontaneously and this inhibits VD4 and inhibits and then excites IP3.
- IP3 firing inhibits VD4 and excites DA1, which produces further inhibition of VD4.
- IP3 stops generating action potentials, which releases VD4 from the inhibition.
- This allows VD4 to generate a series of action potentials, which leads to inhibition of both DA1 and IP3.
- VD4 then stops firing action potentials, which releases DA1 and IP3 from inhibition. 8. DA1 can now spontaneously fire and a new cycle begins.J. There are three general types of things going on here. There are three general types of things that one must know to understand a neural circuit.
- Anatomy; what is connected to what.
- Synaptic effects of the neurotransmitters. 3. The intrinsic properties of the neurons.
a. Intrinsic properties are like the neuron’s personality.
b. Examples of intrinsic properties are whether or not a neuron is spontaneously active.
c. What produces intrinsic properties? The type of voltage-gated ion channels a particular neuron expresses is a major determinant of its intrinsic properties.

Summary of the pond snail CPG

Below is a diagram of a lateral view (side view) of a human brain depicting in simplified form the areas of the cortex involved in these types of motor acts.
- The central sulcus divides the frontal and parietal lobes.
- The primary motor cortex (1  motor cortex) contains neurons that project directly to -motor neurons in the brainstem and spinal cord. The axons of these cortical
neurons form what is known as the pyramidal tract, until they reach the junction of the medulla oblongata and spinal cord. At that point, the axons cross the midline and form what is called the pyramidal decussation and then form the lateral corticospinal tract after crossing (same axons with different names at different points) before synapsing on the  -motor neurons of the ventral horn. - The primary somatosensory cortex (1  somatosensory cortex) is, by definition, the first cortical area receiving synaptic input conveying somatosensory information. (Note that primary visual cortex is the first cortical area receiving visual information).
- The 1  motor cortex and 1  somatosensory cortex are both somatotopically mapped, and these maps are in parallel. There are projections from neurons in 1 somatosensory cortex to neurons in corresponding regions of 1  motor cortex.
- Neurons in the premotor cortex project to neurons in 1  motor cortex.
- Neurons in the posterior parietal cortex project to premotor cortex.

There are three cortical areas that perform different stages in the generation of complex movements… The first is posterior parietal cortex, which projects to the second, which is premotor cortex, which projects to the 3rd, which is 1  motor cortex.
- The neurons in the posterior parietal cortex act as a spatial map, which provides you with information about where you are with respect to a goal object.
- The premotor cortex in involved in planning a complex motor act.
- The 1  motor cortex is more directly involved in directing the execution of the task.
Evidence that posterior parietal cortex acts as a spatial map comes from humans with brain damage of this area.
- Patients with brain damage in the posterior parietal cortex neglect the contralateral half of the body and the contralateral side of objects.
- Patients with brain damage in the posterior parietal cortex also have a dramatic loss of the ability to use spatial maps.
a. They cannot use a road map.
b. They can’t navigate. Even in a bathroom they can get lost (they can’t map where they are relative to the environment they are in).
How do we know that the 1  motor cortex is somatotopically organized?
- If you electrically stimulate different areas of cortex you will find that the 1  motor cortex is the region with the lowest threshold for producing movement.
- When you stimulate, you get movement on the contralateral side of the body. 3. Somatotopic organization…
a. As you move the stimulating electrode across adjacent regions in the 1  motor cortex, you will produce movement in more or less adjacent regions of the body. e.g. the toes, then the foot, the legs……then each of the fingers of a hand, then the face.
b. You will also find that the hands and face have a disproportionally large representation in 1  motor cortex, as compared to other body areas that are larger, such as the trunk.
c. When you stimulate in different areas of the 1  motor cortex, you get movements in different areas of the body on the contralateral side.
How does this map in the 1  motor cortex come about?
- It is the anatomy, what is connected to what, that creates the map in the 1  motor cortex.
- Below is a diagram of a coronal section of a human brain and spinal cord, depicting the anatomical connections that create the somatotopic map.
a. If there was a cut at location 1, the person would have paralysis of the face, hand, and foot on the side of the body contralateral to the cut.
b. If there was a cut at 2, the person would have paralysis of the hand and foot on the side of the body ipsilateral to the cut.
c. If there was a cut at 3, the person would have paralysis of the foot on the side of the body ipsilateral to the cut.
d. If there was brain damage to the 1  motor cortex in the region of the hand, the person would have paralysis of the hand on the side of the body contralateral to the brain damage.
e. These cuts and lesions do not produce total paralysis. The person cannot do complex tasks such as writing, etc. But they can still do thing like walk or climb a tree (talking about monkeys here). Why is this? Because the mesencephalic locomotor nucleus and spinal CPGs are still intact. They are parts of a different neural motor control system, one producing locomotion. So, there are different motor control systems with different functions and they may control the same part of the body [hands write and are involved in locomotion, in the same way the mouths are involved in speaking (one control system) and chewing (generated by a different control system)]. Different control systems may have inputs to the same motor neurons.

What do these neurons in 1  motor cortex do? There are three lines of evidence.
- When you stimulate in the 1  motor cortex you get movement on the contralateral side of the body.
- If you lesion the 1  motor cortex, you lose complex movement on the contralateral side of the body.
- Are action potentials in these neurons actually producing movement? If so, then the action potentials should precede the movement. Below is an experiment that demonstrates this.
a. A group recorded from neurons in monkey 1  motor cortex, and the monkey was taught to twist a bar (thus he is flexing his wrist).
b. They found that neurons in primary motor cortex generate action potentials just before the movement starts, which suggests that the action potentials generated by these neurons in 1  motor cortex are indeed producing the movement.
Evidence that the premotor cortex in involved in planning movement.
- An experiment was done using PET scans. A PET scan gives you an assay of increased activity of collections of neurons in the cortex, so you can determine what part of the brain is active while someone is performing different activities. This is a noninvasive procedure and can be done on humans.
- The subjects were asked to do a simple repetitive movement of the fingers, and it was found that there was increased activity in the 1  motor cortex and 1  somatosensory cortex.
- Next the subjects were asked to perform a more complex, learned sequence of movements with their fingers. It was found that this complex movement also produced increased 1  motor and 1  somatosensory cortex, but there was also an increase in neural activity in the premotor cortex.
- Finally, the subjects were asked to mentally perform a complex, learned sequence of movements of the hands (but not actually do the movements). While doing this “mental rehearsal” it was found that there was increased activity only in the premotor cortex.
- All of this suggests that the premotor cortex is involved in planning movements.
The cerebellum & motor coordination
A. Axons of M1 pyramidal neurons project to MNs of the ventral horn to initiate voluntary movements.
- Proprioceptive sensory feedback from the muscles project through the dorsal root to the spinal cord, where it then projects rostrally to:
a. S1, primary somatosensory cortex b. the cerebellum - S1 relays the sensory information to M1 in a point-to-point fashion (i.e. somatotopically).
Thus, M1 and the cerebellum receive information from the muscles about the outcome of the initiated motor output.
B. Axon collaterals of M1 pyramidal neurons also project to the cerebellum. This is part of the cerebellar motor loop.
- Collaterals of axons from M1 pyramidal neurons project to the cerebellum via the pons of the brainstem.
- The cerebellum projects back to the primary motor cortex via the thalamus. Thus, the cerebellum receives information about the intended voluntary motor output.
The cerebellum then compares what was intended with what has happened, and compensatory modifications are made in cerebellar circuits to more closely match the intended output with the actual outcome of the planned sequence of movements. Thus, the cerebellum is a site of motor coordination and motor learning. Think about this in terms of learning a new skill. Movements that were initially disjointed become, with practice, more and more coordinated until you can perform the activity almost unconsciously.
The somatotopic map in M1 (Looks identical to map in S1)

“See I?
I See, I See”

