Nuclei & Euk Gene Reg (PSET 4) Flashcards
Why can a large nuclear protein get into the nucleus but a smaller version treated with trypsin cannot? (4-2)
The larger protein has a NLS. (Size is not what matters, binding to pores/receptors does).
If the NLS is on the tail of nucleoplasmin (promotes nucleosome assembly) is not needed for function, do you still need the tail after the tail is localized to the nucleus? (4-2)
Yes: After mitosis/meiosis you need a signal to redirect the nuclear proteins back into the reformed nucleus.
Nucleosomes (4-3)
DNA in chromatin is in nucleosomes. A nucleosome ~145 BP DNA wrapped around histone (core) and linker region ~55 BP. This structure protects the DNA from cleavage or crosslinking (psoralen) enzymes.
Where are nucleosomes found? (4-3)
Nucleosomes are usually found throughout the transcribed regions, including right ahead of and right behind RNA polymerase. Nucleosomes are probably a little looser than in untranscribed regions, and the single nucleosome where the RNA pol is may be partially unfolded or absent (temporarily), but there is no wholesale loss of nucleosomes, though some regulatory regions may lose nucleosomes.
How do transcription factors affect nucleosomes? (4-3)
Tissue specific transcription factors bind to regulatory regions (enhancers) that are usually (but not always upstream of the genes they regulate. The binding of the transcription factors displaces nucleosomes, exposing the DNA to DNase or psoralen. So DNA in enhancer region of an actively transcribed geen should be very sensitive to DNase and should give a random cross-linking pattern with psoralen (psoralen enzymes crosslink DNA)
In a strand of DNA with 4 nucleosomes, how many do you have of each of the following? (4-4A) i) molecules of DNA? ii) base pairs iii) Molecules of H2A
i) One continuous DNA molecule. ii) Each of the 4 histones contains ~200 bps DNA: 4 x 200 = 800 bps long iii) Each nucleosome core contains 8 histone molecules: 2 each of H2A, H2B, H3, and H4. So, 2 molecules H2A in each nucleosome x 4 nucleosomes = 8 molecules H2A
What is the difference between mitotic chromatin and chromatin in Interphase? (4-4B)
Mitotic chromatin should be shorter because it is more tightly folded than “beads on a string” as in interphase. Histone H1 is required for higher orders of folding (there is one H1 per nucleosome). The number of nucleosomes per stretch of DNA does not change but the higher orders of folding are different: the chain of nucleosomes is folded back on itself and the folded chromatin is folded back on itself again several times, yielding a thicker. shorter structure.
Describe the chromatin in transcribed regions of genes. (4-4C)
The section must contain nucleosomes during interphase. Cis (non transcribed) regulatory regions of active genes generally do not have nucleosomes – the DNA is bound to proteins but the proteins = transcription factors or co-activators instead of histones. This includes the promoter, enhancer, and proximal regulatory regions. It may displace one nucleosome at a time, but doesn’t remove them in wholesale fashion. Both exons and introns are transcribed, so the region could be from either one.
Suppose a cell increases rate of ribosomal production. How is the rate of protein synthesis inside the nucleus change? How does the rate of rib proteins moving INTO of the nucleus change? How does the rate of rib proteins moving INTO the nucleus change? (4-7)
Protein synthesis inside the nucleus should STAY THE SAME AT ZERO. Rate of individual ribosomal proteins moving out of nucleus should STAY THE SAME AT ZERO. (Rate of subunits moving out of the nucleus would increase but ribosomal proteins do not move out of the nucleus as separate molecules.) Rate of proteins moving into the nucleus should increase. (as proteins move in to meet rRNA)
What happens to # of nucleosomes and chromatin width from M to G1? (4-7)
of nucleosomes: stays the same chromatin width: decreases (as the chromatin de-condenses it gets longer and thinner)
What happens to lamins during… (i) G2 to M? (ii) G1 to S? (4-7)
(i) During G2 to M: lamin depolymerization increases (ii) During G1 to S: lamin depolymerization should stay the same at zero
What is the structure of lamins? (4-7)
Lamins are IF and have a NLS.
Do you expect E. coli to use similar gene regulation involving histones as we use? (4-8)
Prokaryotes do not have histones.
Where do you find histones? (4-9)
Each nucleosome contains an octamer of 8 molecules of histone: 2 each of H2A, H2B, H3, and H4. And 1 molecule of H1. Spread out about every 200 bps of DNA. H1 is found at “linker regions”, connecting two cores. (Note: You can reconstitute nucleosomes in vitro without H1.)
How do you describe a blot pattern of chromatin cut with nuclease that has multiple bands? (4-9)
Nuclease must have cut at random (since bands are different lengths, each band cannot represent the DNA from a single nucleosome. Instead each band must represent a multiple of 200 bps or the DNA from one or more nucleosomes). The nuclease cuts in between nucleosome cores, in the linkers, but it does NOT cut a EVERY linker. Thus, the region that hybridizes to the probe can be a fragment of 200 bps, 400, 600…
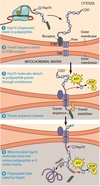
Experiment Showing That Polypeptides Span
Both Mitochondrial Membranes During Import.
To demonstrate
that polypeptides being imported into mitochondria span both membranes
at the same time, a cell-free import system was incubated on
ice instead of at the usual temperature of 37°C. At low temperature,
polypeptides start to enter the mitochondrion, but their translocation
soon stalls. Under these conditions, the transit sequence is cleaved by
transit peptidase in the matrix, indicating that the N-terminus of the
polypeptide is inside the mitochondrion. At the same time, most of
the polypeptide chain is accessible to attack by proteolytic enzymes
added to the outside of the mitochondrion. This means that the
polypeptide must span both membranes during import, presumably
at a contact site between the two membranes.

Posttranslational Import of Polypeptides
into the Mitochondrion.
Like cotranslational import into the ER,
posttranslational import into a mitochondrion involves a signal
sequence (called a transit sequence in this case), a membrane receptor,
pore-forming membrane proteins, and a peptidase. However, in the
mitochondrion, the membrane receptor recognizes the signal
sequence directly, without the intervention of a cytosolic SRP. Furthermore,
chaperone proteins play several crucial roles in the
mitochondrial process: They keep the polypeptide partially unfolded
after synthesis in the cytosol so that binding of the transit sequence
and translocation can occur (steps – ); they drive the translocation
itself by binding to and releasing from the polypeptide within
the matrix, which is an ATP-requiring process (step ); and they
help the polypeptide fold into its final conformation (step ). The
chaperones included here are cytosolic and mitochondrial versions
of Hsp70 (light blue) and a mitochondrial Hsp60 (not illustrated)